Deficiencies in thiamine have resulted in numerous deleterious effects observed in both people and the environment. Most notably, deficiencies are responsible for the Wernicke-Korsakoff syndrome and Beriberi disease prevalent in alcoholics in developed nations and undernourished populations in developing nations. These conditions primarily yield neurological and cardiac symptoms that can range from dysfunction to death. Deficiencies are also implicated in other conditions including diabetes, depression, Alzheimer’s disease, and dementia.
In an ecological context, thiamine deficiencies result in muscle weakness, inability to spawn, and poor survival rate of fry from predator fish including lake trout and Atlantic salmon. Such deficiencies have also affected alligators, wild birds, as well as various species of livestock.1-4 Its limited availability, yet physiologic necessity, has yielded competition amongst numerous organisms.
In many of these conditions, administration of high doses of thiamine can be curative, while continued lack of thiamine can yield irreparable harm, hence early recognition of deficiency is imperative.
Salmonid species of fish, including Atlantic salmon, chinook salmon, coho salmon, and lake trout, are especially affected by thiamine deficiency. Certain species of forage fish, most notably alewifes and rainbow smelt, are known to have high levels of thiaminase enzymes. As these are prey fish for salmonids, although the dietary intake of thiamine by the latter may be adequate, thiaminase enzymes released upon digestion cause the breakdown of thiamine in the gut before it can be absorbed. Thus, maternal thiamine levels are often deficient causing inability to spawn, reduced predator-prey instinct, and poor survival rates of their offspring. This not only has a notable impact in the ecological realm, but also an economic loss to the sport-fishing industry.3 Marked losses of salmonids have been noted in hatcheries, for example, causing 60-90% mortalities in coho salmon near the Great Lakes.22 Several conditions are associated with thiamine deficiency in fish, including the M74 Syndrome, primarily affecting Atlantic salmon in the Baltic Sea; the Early Mortality Syndrome (EMS), affecting salmonids from the Great Lakes; and the Cayuga Syndrome, affecting Atlantic salmon in the NY Finger Lakes.4,5,23,26,27
Other aquatic species are also impacted by thiamine deficiency. In the Baltic Sea area, wild birds including herring gulls and eiders have suffered from a paralytic syndrome resulting in their population decline. The progression of this syndrome includes the inability to keep wings folded, loss of walking ability and flight, seizures, total paralysis, and death. In a subset of herring gulls, recovery of paralyzed adult birds was possible by treatment with injected thiamine.1 Aside from Atlantic salmon and common eiders, a recent study has shown widespread deficiency in the Northern hemisphere in European eel, American eel, sea trout, and blue mussel.25 This has been evidenced not only by decreased thiamine levels and TDP-dependent enzyme activity, but also sublethal viability and behavioral abnormalities. This study took place at 45 stations in 15 regions ranging from Maryland to Iceland to Stockholm, suggesting that environmental thiamine deficiency is more widespread and severe than previously realized.
Impact in the Veterinary realm
Livestock Health
In livestock, the clinical term associated with the effects of thiamine deficiency is polioencephalomalacia or PEM. This condition encompasses symptoms including loss of appetite, reduced heart rate, convulsions, and death. A characteristic symptom of thiamine deficiency in livestock is 'star-gazing' where paralysis of the neck muscles causes the animal to look upward.1 Most commonly, PEM is found in cattle, goats, and sheep that consume thiaminase-containing plants (such as bracken fern, horsetail, and nardoo fern); consume high levels of sulfur-containing foods; have an abnormally high grain intake; or are undergoing treatments to control bacterial or parasitic infections.
Many bacteria are capable of thiamine synthesis and those present within the rumen of these mammals can contribute positively to their thiamine status. For example, in one study carried out on sheep, despite a low dietary intake of ≤0.3 mg thiamine, between 1.44 and 3.23 mg/day produced through endogenous microbial sources was found to enter the small intestine.6 However, when dietary changes or antibiotics disrupt the normal composition of the gut microflora, this relatively 'endogenous' source of thiamine is lost. For example, diets high in grains often are a good source of thiamine, but also reduce the pH of the rumen and lend towards the growth of thiaminase-containing bacteria and reduction of growth of the normal microflora.
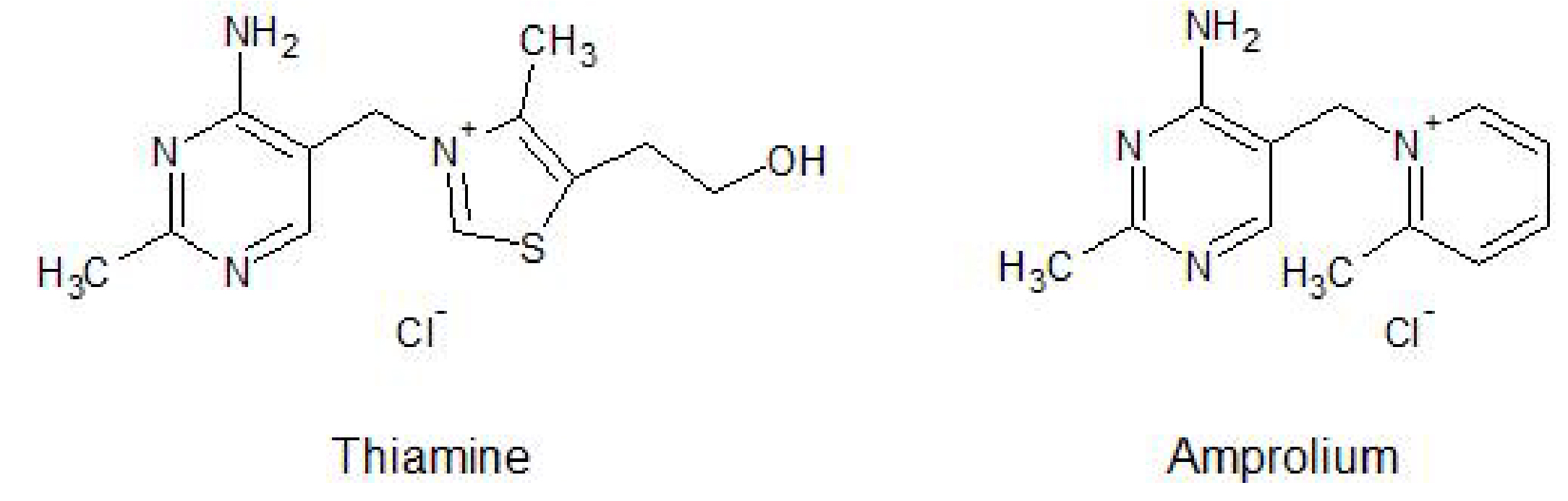
Further, certain anti-parasitic agents, most notably amprolium which shares the aminomethylpyrimidine ring and cationic nitrogen of thiamine (Figure 1), can block the uptake of thiamine from the gut by acting as a competitor for its transport.
Figure 1. Chemical structures of thiamine versus amprolium, a therapeutic agent used for parasite control in poultry and livestock
In fact, inhibition of thiamine uptake is the mechanism of action of amprolium against parasites - certain coccidia protozoa, such as Eimeria bovis and Eimeria zurnii, have a high requirement for thiamine and thus are more susceptible to thiamine deficiency than the host. However, upon prolonged treatment or in animals with dietary deficiencies otherwise, symptoms of thiamine deficiency can result.7,8,9
From 2009-2017, several voluntary recalls of cat food due to low thiamine levels have been announced through the FDA. The recalls have included dry, canned, and raw frozen food from well-known manufacturers including Purina, Iams, Wellness, Primal Pet Foods, 9-Lives, and Diamond.20 Cats are particularly susceptible to thiamine deficiency due to their relatively demanding nutritional requirement. For example, they require 1.4 mg thiamine per 1000 kcal consumed, whereas dogs require only 0.56 mg thiamine per 1000 kcal consumed.21,22
As with food fit for human consumption, thiamine is often added to pet food to balance the losses that occur during processing upon heating and storage. In particular, in canned food, a sterilization process is carried out to eradicate pathogenic bacteria, however, high temperatures and the potentially alkaline conditions present in gelling agents are prime conditions for the destruction of thiamine.9 The losses due to cooking can be further exacerbated in home-prepared pet foods, the processing of which may be variable and with no monitoring of the losses of thiamine available. Further, while the use of sulfites in meat products for human consumption is banned in many countries, the same is not necessarily extended to pet foods and can contribute to the breakdown of thiamine.9,23
Additionally, in fish-containing foods, there may be seasonal variation in the type of fish caught, and thus the thiaminase content of the foods.23 While this is typically inactivated during the canning process, the processing of foods in homemade diets may not be adequate. Ingestion of raw fish where thiaminases have not been inactivated has yielded thiamine deficiency not only in cats, but also dogs and reptiles.
Thiamine defiencies in cats also occur through routine feeding of foods intended for supplemental use or dog food which are not nutritionally balanced to meet the long-term health needs of cats.9
In 1932, in a population of more than 200 captive foxes, one third perished within a one month period. Chastek's paralysis, termed for the farm in Minnesota where the condition was first observed, encompasses symptoms including anorexia, weakness, staggering, loss of control of body movements (ataxia), and ultimately death.10 The association with thiamine deficiency was made in 1936 through a change in diet to raw fish during the autumn months and ability to reproduce the symptoms in foxes with thiamine deficient diets experimentally. This condition was observed in both mink and foxes fed a diet of fresh frozen smelt (Thaleichythys pacificus) with symptoms ameliorated by thiamine injections and return to a non-fish diet within a short period.11
In captive marine mammals, including sea lions, bottlenose dolphin, grey seals, and killer whales, thiamine deficiency has been associated with diets composed of herring (Clupea harengus) and smelt (Osmerus mordax).12
Figure 2. Examples of thiamine deficiency on animal populations throughout history
Impact in the clinical realm
Thiamine deficiencies have plagued man throughout history (Figure 3), exemplified by the widespread outbreaks of Beriberi disease detailed previously. However, Beriberi is not the only thiamine deficiency related disease. In 1881, the German professor Carl Wernicke observed an acute change in mental status, opthalmic symptoms, and loss of control of body movements (ataxia) in three patients with a history of alcohol abuse or poisoning. These patients had similar brain lesions found upon autopsy and culminated in the coining of the condition as 'Wernicke's disease' or 'hemorrhagic polioencephalitis.' 13-15 Shortly thereafter, in 1887, a Russian professor S.S. Korsakoff noted chronic symptoms of amnesia along with structural brain lesions in primarily alcoholic patients, terming the condition 'psychosis polyneuritica' or 'Korsakoff's psychosis'. These two conditions were later associated as a progressive disease termed the Wernicke-Korsakoff syndrome, which is associated with thiamine deficiency due to poor diet and imparied uptake from the GI tract. It commonly affects chronic alcoholics, but also those with restricted diets or capacity for thiamine absorption from foods, such as those with anorexia nervosa or who have undergone gastric bypass surgery. The symptoms of the acute phase ('Wernicke syndrome') which occur in the earlier stages can be ameliorated with thiamine supplementation, whereas permanent damage can result from the chronic phase (Korsakoff syndrome).
Even in recent years, as has been the case with cat foods, thiamine deficiency in commercial foods fit for human consumption has contributed to morbidity and mortality. In 2003, a soy-based infant formula defectively manufactured without the addition of thiamine was distributed in Israel and caused symptoms of thiamine deficiency in 20 infants leading to 4 deaths.16,17 Similar to the issues experienced in the Japanese navy more than 100 years earlier, in 2005, 15 cases of thiamine deficiency leading to 2 deaths on a commercial fishing vessel in Thailand resulted and were attributed to fish and rice diets.18 Despite the growing body of knowledge, as recently as 2011, an outbreak due to dietary insufficiency occurred in school-aged children in Bhutan, leading to 31 cases and 2 deaths.19
Figure 3. Examples of thiamine deficiency on human populations throughout history
A variety of other conditions are attributed to thiamine deficiency in people, including African Seasonal Ataxia, Konzo, and Tropical Ataxic Neuropathy.
Other conditions are not caused by thiamine deficiency, but rather by a lack of or dysfunction of transporters responsible for thiamine transport, enzymes needed for conversion to TDP, or those relying on TDP for function. For example, thiamine-responsive megaloblastic anemia (TRMA, or Roger's syndrome) is a genetic condition where the thiamine transporter (ThTR1) is deficient, which restricts thiamine uptake. In maple syrup urine disease (MSUD), the enzymes involved in the branched chain alpha-keto acid dehydrogenase complex are genetically deficient. Deficiencies in these enzymes result in the build-up of isoleucine, leucine, and valine, leading to toxic effects. As these enzymes use TDP as a co-factor, in patients with some level of expression, the limited available enzyme can be stimulated by high doses of thiamine which can ameliorate symptoms.
A summary of the conditions associated with thiamine deficiencies and their causes is provided in Table 1.
Table 1. Conditions resulting from thiamine deficiency or related causes in people and animals
Disease/Condition
Species affected
Predominant symptoms
Common cause
Ref.
Conditions affecting people
Amish lethal microcephaly
Humans
Small head size, elevated urinary levels of 2-ketoglutaric acid, and death within the first year
Genetic lack of thiamine transporter (impaired absorption of thiamine)
1
African Seasonal Ataxia
Humans
Impaired balance (acute onset)
Diet of Anaphe venata larvae (thiaminases)
2,3
Biotin-responsive basal ganglia disease
Humans
Involuntary muscle contractions, confusion, coma, death
Genetic lack of thiamine transporter (impaired absorption of thiamine)
4
Dry beriberi
Humans
Peripheral neuritis, muscle weakness, loss of sensation
Cardiac or neurologic symptoms as described for wet/dry beriberi
Inadequate maternal thiamine intake
6
Konzo
Humans
Paralysis of lower limbs
Diet of cassava (destruction of thiamine by cyanogenic glycosides)
7
Tropical ataxic neuropathy
Humans
Optical and auditory neuropathy
Diet of cassava, rice, and alcohol consumption (destruction of thiamine, nutritional deficits, impaired absorption)
8
Korsakoff
Humans
Loss of recent memory, coma, death
Chronic alcoholism (impaired absorption of thiamine)
5
Maple syrup urine disease
Humans
Impaired mental development, seizures, coma, death
Genetic lack of branched chain ketoacid dehydrogenase (impaired conversion by TDP-dependent enzyme)
9
Thiamine-responsive megaloblastic anemia
Humans
Hearing loss, diabetes, optical impairments
Genetic lack of thiamine transporter (impaired absorption of thiamine)
10
Wernicke
Humans
Optical impairments, confusion, delirium, death
Chronic alcoholism (impaired absorption of thiamine)
5
Conditions affecting other species
Cayuga Syndrome
Landlocked Atlantic salmon
Yolk-sac opacities, hydrocephalus, marked mortality rate of fry
Diet of alewives/smelt (thiaminases)
11
Chastek’s paralysis
Fox and mink
Anorexia, loss of motor control, death
Diet of raw fish (thiaminases)
12
Early Mortality Syndrome
Salmonids
Swimming in spirals, anorexia, lethargy, hyperexcitability, hydrocephalus, marked mortality rate of fry
Diet of alewives/smelt (thiaminases)
13
M74 Syndrome
Atlantic salmon
Impaired development of fry; Neurological, muscular symptoms
Diet of alewives/smelt (thiaminases)
14
Polioencephalomalacia
Cattle, goats, deer, camelids, and sheep
‘Star-gazing’, anorexia, loss of motor control, coma, death
Diet of high grain content/thiaminase-containing plants (also high sulfur intake)
15
Polyneuritis
Chicken, turkey, pigeons
Anorexia, paralysis, ‘Star-gazing’, death
Dietary insufficiency, treatment with amprolium
16
Biochemical ramifications of thiamine deficiency
While dietary insufficiency or breakdown of thiamine by thiaminases is a clear cause of thiamine deficiency, what bears further mention are the biochemical mechanisms that underlie the outward ramifications. As discussed on the biochemistry page, thiamine in its diphosphate form is a cofactor for several enzymes involved in carbohydrate metabolism. If an organism is thiamine deficient, these enzymes, and the pathways that they are involved in, are hindered. This limits the production of ATP, ribose-5-phosphate, and NADPH which originate via the citric acid cycle and pentose-phosphate pathways, respectively.
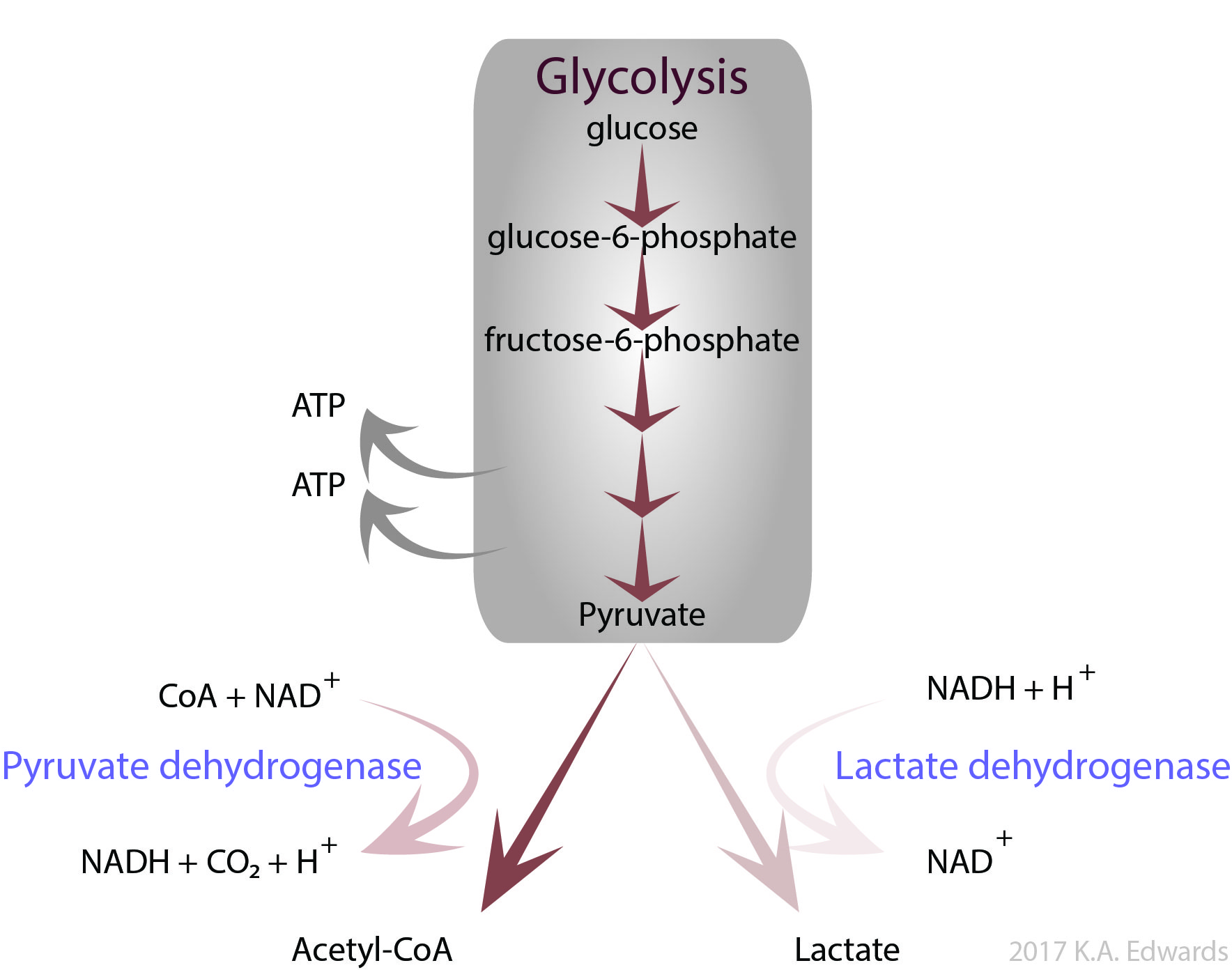
In the case of high carbohydrate intake but low thiamine, while there is plenty of glucose to initiate these pathways, the limiting factor is then functional enzymes to process the downstream intermediates. For example, the glycolysis pathway produces pyruvate from glucose (Figure 4). Pyruvate is converted to acetyl-CoA by pyruvate dehydrogenase (PDH), a TDP-dependent enzyme, prior to entering the citric acid cycle. Under conditions of low oxygen, or if an organism is deficient in thiamine, pyruvate accumulates and is instead converted to lactate by lactate dehydrogenase.
Figure 4. Pyruvate formed from glycolysis is converted via pyruvate dehydrogenase to acetyl CoA which enters the citric acid cycle or by lactate dehydrogenase to lactate under anerobic conditions, respectively
Build up of lactate is normal in muscles during exercise, but chronic generation of lactate can lead to lactic acidosis. This condition yields a low physiological pH and results in nausea, rapid breathing, and muscle weakness. These same symptoms can occur if there is a deficiency in the PDH enzyme itself. Rare genetic abnormalities result in PDH either being poorly expressed or expressed in a disfunctional form, resulting in overall low ATP generation. As the brain requires large amounts of energy, it is markedly impacted by PDH deficiencies and patients have marked neurological impairments, usually from birth. These impairments include small head size (microcephaly). PDH deficiency is often treated with high doses of thiamine to ensure optimal function of available PDH, although not all cases are thiamine-responsive. If the cause of PDH deficiency is mutations that lower the affinity of the enzyme towards TDP, high doses of thiamine can help to ameliorate this condition.24
As another example, TDP is the cofactor involved in breakdown of branched chain amino acids (leucine, isoleucine, and valine). These amino acids are absorbed upon digestion of proteins from the diet or are present due to normal breakdown of proteins in the body. Branched chain alpha-ketoaciddehydrogenase (BCKD) is responsible for conversion of these amino acids to their thioesters with coenzyme A, which are intermediates in the formation of acetyl CoA, succinyl CoA, and acetoacetate (Figure 5).
Figure 5. Breakdown of branched chain amino acids (leucine, valine, and isoleucine) by BCKD
Acetoacetate is a precursor to Acetyl CoA; acetyl CoA is a starting material for the citric acid cycle; and succinyl CoA is an intermediate of the citric acid cycle. If thiamine is deficient, or there is a defect in BCKD, not only is the energy available from the citric acid cycle limited, but the breakdown of these amino acids is impaired. In a genetic condition known as Maple Syrup Urine Disease (MSUD), defective BCKD causes an accumulation of these amino acids which are excreted in the urine. Unfortunately, patients with this rare disease typically have severe developmental retardation.
References
1. Balk L, Hägerroth P-Å, Åkerman G, et al. Wild birds of declining European species are dying from a thiamine deficiency syndrome. Proceedings of the National Academy of Sciences. 2009;106(29):12001-12006.
2. Ross JP, Honeyfield DC, Brown SB, et al. Gizzard shad thiaminase activity and its effect on the thiamine status of captive American alligators Alligator mississippiensis. Journal of Aquatic Animal Health. 2009;21(4):239-248.
3. Carvalho PS, Tillitt DE, Zajicek JL, et al. Thiamine deficiency effects on the vision and foraging ability of lake trout fry. Journal of Aquatic Animal Health. 2009;21(4):315-325.
5. Keinänen M, Uddström A, Mikkonen J, et al. The thiamine deficiency syndrome M74, a reproductive disorder of Atlantic salmon (Salmo salar) feeding in the Baltic Sea, is related to the fat and thiamine content of prey fish. ICES Journal of Marine Science: Journal du Conseil. 2012;69(4):516-528.
6. Breves G, Hoeller H, Harmeyer J, Martens H. Thiamin balance in the gastrointestinal tract of sheep. J Anim Sci. 1980;51(5):1177-1181.
7. Loew FM, Dunlop RH. Induction of thiamine inadequacy and polioencephalomalacia in adult sheep with amprolium. American Journal of Veterinary Research. 1972;33(11):2195-2205.
8. Davis J, Gookin J. Antiprotozoan Drugs. In: Riviere J, Papich M, eds. Veterinary Pharmacology and Therapeutics. 9th ed: John Wiley & Sons; 2009.
9. Markovich JE, Heinze CR, Freeman LM. Thiamine deficiency in dogs and cats. Journal of the American Veterinary Medical Association. 2013;243(5):649-656.
10. Evans CA, Carlosn WE, Green RG. The pathology of Chastek Paraysis in Foxes. Am J Pathol 1942; 18(1): 79-91.
11. Long JB, Shaw JN. Chastek paralysis produced in Oregon mink and foxes by feeding fresh frozen smelt. N Am Vet 1943; 24: 234-237.
12. Geraci JR. Diet-induced thiamine deficiency in captive marine mammals. http://www.vin.com/apputil/content/defaultadv1.aspx?pld=11081&meta=Generic&catid28893&id=3979809&print=1
13. Goodwin DW. The wernicke-korsakoff syndrome: A clinical and pathological study of 245 patients, 82 with post-mortem examinations. JAMA. 1972;219(3):389-389.
15. Goodwin DW. The wernicke-korsakoff syndrome: A clinical and pathological study of 245 patients, 82 with post-mortem examinations. JAMA. 1972;219(3):389-389.
16. Fattal-Valevski A, Kesler A, Sela B-A, et al. Outbreak of Life-Threatening Thiamine Deficiency in Infants in Israel Caused by a Defective Soy-Based Formula. Pediatrics. 2005;115(2):e233-e238.
17. Shamir R. Thiamine-deficient infant formula: what happened and what have we learned? Ann Nutr Metab. 2012;60(3):185-187.
18. Doung-ngern P, Kesornsukhon S, Kanlayanaphotporn J, Wanadurongwan S, Songchitsomboon S. Beriberi outbreak among commercial fishermen, Thailand 2005. Southeast Asian Journal of Tropical Medicine & Public Health. 2007;38(1):130-135.
23. Loew FM, Martin CL, Dunlop RH, Mapletoft RJ, Smith SI, Naturally-occurring and experimemental thiamin deficiency in cats receiving commercial cat food, Can Vet J. 1970; 11(6): 109-113.
24. Naito E, Ito M, Yokota I, et al. Thiamine-responsive pyruvate dehydrogenase deficiency in two patients caused by a point mutation (F205L and L216F) within the thiamine pyrophosphate binding region.Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease. 2002;1588(1):79-84.
27. Fitzsimmons JD, Brown SB, Honeyfield DC, Hnath JG. A review of Early Mortality Syndrome (EMS) in Great Lakes Salmonids: Relationship with Thiamine Deficiency, AMBIO, 1999; 28(1): 9-15.
Table 1 references:
Lindhurst MJ, Biesecker LG. GeneReviews® [Internet]. In: Pagon RA AM, Ardinger HH, et al., ed. Amish Lethal Microcephaly: University of Washington, Seattle; 2003. (https://www.ncbi.nlm.nih.gov/books/NBK1365/)
Nishimune T, Watanabe Y, Okazaki H, Akai H. Thiamin is decomposed due to Anaphe spp. entomophagy in seasonal ataxia patients in Nigeria. The Journal of nutrition. 2000;130(6):1625-1628.
McCandless D, African seasonal ataxia, in Thiamine Deficiency and Associated Clinical Disorders, Springer
Shamir R. Thiamine-deficient infant formula: what happened and what have we learned? Ann Nutr Metab. 2012;60(3):185-187.
Adamolekun B. Etiology of Konzo, epidemic spastic paraparesis associated with cyanogenic glycosides in cassava: role of thiamine deficiency? Journal of the Neurological Sciences. 2010;296(1-2):30-33.
Adamolekun B. Thiamine deficiency and the etiology of tropical ataxic neuropathy.Int Health2010; 2(1): 17-21.
Neufeld EJ, Fleming JC, Tartaglini E, Steinkamp MP. Thiamine-responsive megaloblastic anemia syndrome: a disorder of high-affinity thiamine transport. Blood Cells Mol Dis. Jan-Feb 2001;27(1):135-138.
Fisher JP, Spitsbergen JM, Iamonte T, Little EE, Delornay A, Pathological and behavioral manifestations of the “Cayuga Syndrome,” a thiamine deficiency in larval landlocked Atlanic salmon, Amer Fish Soc 1995: 7(4): 269-283.
Evans CA, Carlosn WE, Green RG. The pathology of Chastek Paraysis in Foxes. Am J Pathol1942; 18(1): 79-91.