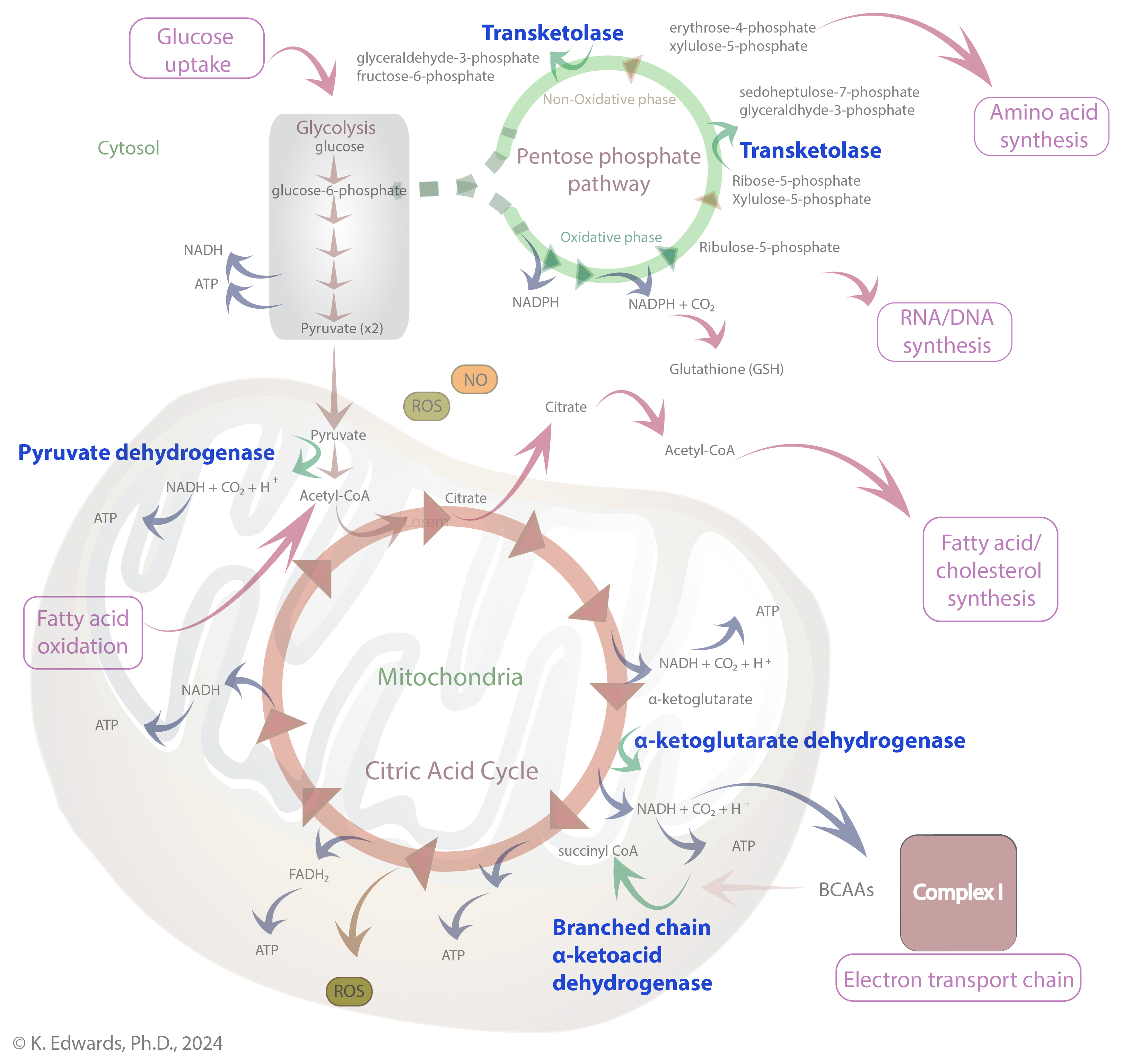
Thiamine (vitamin B1) is essential to the health of all living organisms. In its diphosphate form (also known as TDP, thiamine pyrophosphate, TPP, or cocarboxylase), it serves as a cofactor for enzymes involved in carbohydrate metabolism, including transketolase, α-ketoglutarate dehydrogenase, pyruvate dehydrogenase, and branched chain α-keto acid dehydrogenase. These enzymes are involved in pathways that allow for the production of ATP, NADPH, and ribose-5-phosphate which are critical for generating cellular energy and downstream production of amino acids, nucleic acids, and fatty acids (Figure 1).
TDP is a cofactor for enzymes involved in carbohydrate metabolism, including transketolase, α-ketoglutarate dehydrogenase, pyruvate dehydrogenase, and branched chain α-keto acid dehydrogenase. These enzymes are involved in the link between glycolysis and the citric acid cycle; the citric acid cycle itself; the pentose-phosphate pathway; that allow for the production of ATP, NADPH, and ribose-5-phosphate which are critical for generating cellular energy and downstream production of amino acids, nucleic acids, and fatty acids (Figure 1).
Figure 1. Pathways relying on thiamine diphosphate (TDP)-dependent enzymes. Enzymes relying on TDP as a cofactor are in blue.
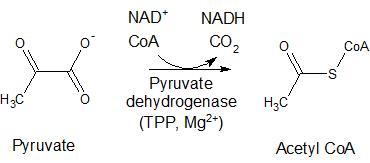
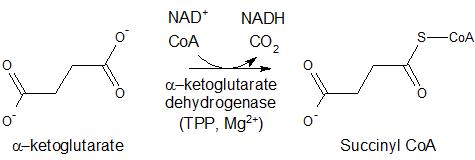
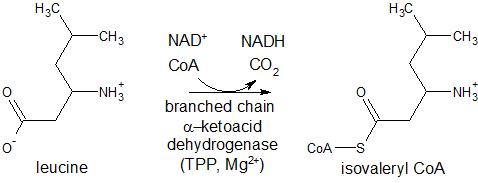
α-ketoglutarate dehydrogenase (KGDH), pyruvate dehydrogenase (PDH), and branched chain α-keto acid dehydrogenase (BCKDC) are found in the mitochondria and serve to oxidize their respective carboxylated substrates. With the loss of CO2, these enzymes form thioesters from these substrates with the concomittant reduction of nicotinamide adenine dinucleotide (NAD+) to form NADH (Figure 2). In the presence of coenzyme A (CoA), PDH oxidizes pyruvate to acetyl coenzyme A (acetyl CoA) which is the starting material for the citric acid cycle; α-ketoglutarate dehydrogenase catalyzes the oxidation of α-ketoglutarate to form succinyl CoA, which is a key intermediate in the citric acid cycle; and BCKDC catalyzes the oxidation of branched chain amino acids, including valine, isoleucine, and leucine, to form the respective thioesters with acetyl CoA. PDH acts similarly in plants and some bacteria for the generation of acetyl CoA. In plants and bacteria, BCDKC is involved in the formation of branched chain amino acid derivatives of long chain hydrocarbons and long-chain fatty acids, respectively.
Figure 2. Reactions of enzymes utilizing TPP as a cofactor, including pyruvate dehydrogenase, alpha-ketoglutarate dehydrogenase, and branched chain alpha-ketoacid dehydrogenase.
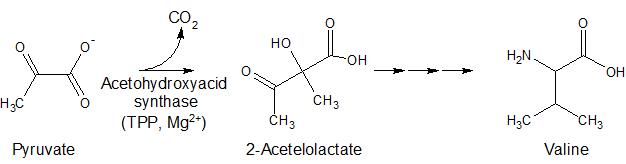
While BCDKC is involed in the breakdown of branched chain amino acids, another TDP-dependent enzyme is involved in their synthesis. In plants and microorganisms, the formation of branched chain amino acids from pyruvate is catalyzed by acetohydroxyacidsynthase (AHAS) (Figure 3).
Figure 3. Formation of branched chain amino acids via acetohydroxyacid synthase
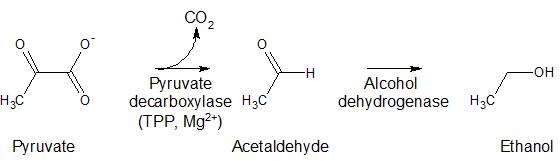
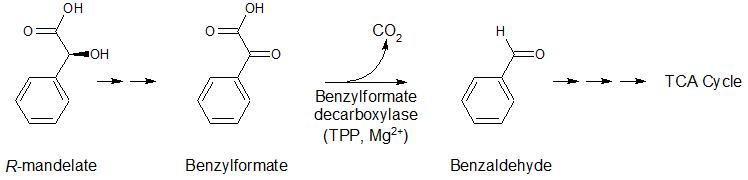
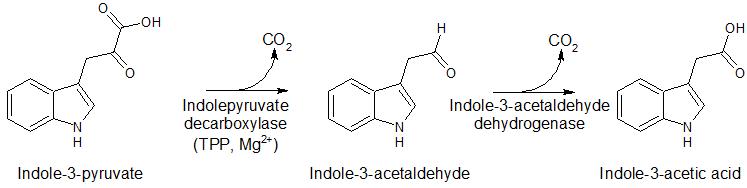
Other enzymes in non-mammalian systems that utilize TDP as a cofactor include alpha-keto decarboxylases (pyruvate decarboxylase (PDC), benzoylformate decarboxylase (BFDC) and indolepyruvate decarboxylase (IPDC)) (Figure 4). These enzymes are involved in the fermentation pathway forming ethanol in yeast; the mandelate pathway allowing bacteria to use mandelic acid as a carbon source; and formation of indole-3-acetic acid, a hormone important for plant development and growth, respectively.
Figure 4. Reactions of enzymes utilizing TPP as a cofactor, including pyruvate decarboxylase, benzoylformate decarboxylase, and indolepyruvate decarboxylase.
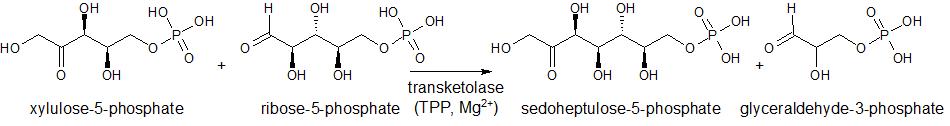
Aside from enzymes that utilize TPP as a cofactor to oxidize their substrates, transketolase enzymes which catalyze the transfer of a 2-carbon glycoaldehyde fragment also use TPP (Figure 5).
Figure 5. Transketolase transfer of a 2-carbon glycoaldehyde fragment from a ketose which contains a single ketone group(xylulose-5-phosphate (donor)) to an aldose which contains a single aldehyde group (ribose-5-phosphate (acceptor)).
In the oxidative phase of the pentose-phosphate pathway, transketolase serves to catalyze the transfer of a glycoaldehyde from xylulose-5-phosphate to ribose-5-phosphate, forming sedoheptulose-7-phosphate and glyceraldehyde-3-phosphate in the process (Figure 1 and Figure 5). In the non-oxidative phase, transketolase catalyzes the transfer of this same fragment, but instead from xylulose-5-phosphate to erythrose-4-phosphate, forming glyceraldehyde-3-phosphate and fructose-6-phosphate in the process (Figure 1).
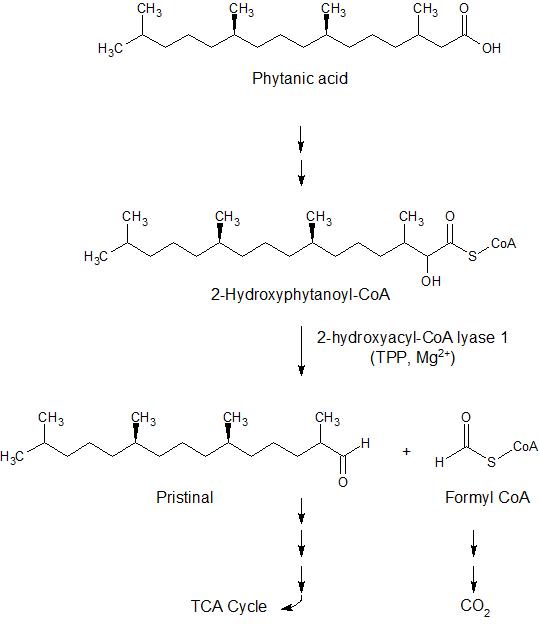
Another enzyme which utilizes TDP as a cofactor is 2-hydroxyacyl-CoA lyase 1 (HACL1).4 Unlike transketolase, PDH, KGDH, and BCKD enzymes which are located in the mitochondria, this enzyme is found in the peroxisomes of mammalian cells. It serves to break down phytanic acid, a branched chain fatty acid which is found in fats present in goat, cow and sheep milk; meats including beef and lamb; and fish.6 In a series of steps, phytanic acid is converted to pristanal while forming formyl CoA (Figure 6). These components can be further oxidized to form CoA derivatives that can enter the citric acid cycle and CO2, respectively. Additionally, HACL1 can act on 2-hydroxy long chain fatty acids in a similar manner, yielding the (n-1) aldehyde of the original fatty acid.
Figure 6. Breakdown of phytanic acid via alpha oxidation to pristanal via HACL1